জোফোবাস মোরিও হল একধরনের অন্ধকারের পোকা যাদের শূককীট সাধারণ নামে সুপারওয়ার্ম, কিংওয়ার্ম, মোরিও ওয়ার্ম বা সহজভাবে জোফোবাস নামে পরিচিত। এটি এমন এক প্রকার পোকা-মাকড়ের শূককীট যা প্লাস্টিক খেয়ে থাকে। অস্ট্রেলিয়ান বিজ্ঞানীরা জোফোবাস মোরিও নামের এই প্রজাতির পোকা খুঁজে পান। এটি মহাপোকা (সুপারওয়ার্ম) হিসাবে পরিচিত। পলিস্টাইরিন বা প্লাস্টিক জাতীয় খাদ্য খেয়ে বেঁচে থাকতে পারে এই পোকাগুলো। গবেষকদদের মতে বিটল শূককীট একটি অন্ত্রের উৎসেচকের মাধ্যমে প্লাস্টিক হজম করে। শূককীটগুলি পূর্ণ আকারে প্রায় ৫০ থেকে ৬০ মিলিমিটার (১.৭-২.২৫ ইঞ্চি) লম্বা হয়।
বিজ্ঞানী ডা. ক্রিস রিঙ্কের মতে মহাপোকাগুলি অতিক্ষুদ্র পুনঃচক্রায়ন কারখানার (মিনি রিসাইক্লিং প্ল্যান্ট) মতো তাদের মুখ দিয়ে পলিস্টাইরিন টুকরো টুকরো করে এবং তারপরে তাদের অন্ত্রে থাকা ব্যাকটেরিয়াকে সেগুলো খাওয়ায়।
ইউনিভার্সিটি অফ কুইন্সল্যান্ড একটি দল তিন সপ্তাহ ধরে মহাপোকার তিনটি দলকে বিভিন্ন ধরনের খাবার খাওয়ায়। যে শ্রেণীটি পলিস্টাইরিন বা প্লাস্টিক জাতীয় খাবার খেয়েছিল তার ওজনও বেড়েছে।
মহাপোকার অন্ত্রে থাকা বেশ কয়েকটি উৎসেচক পলিস্টাইরিন এবং স্টাইরিনকে হজম করতে পারে। আন্তর্জাতিকভাবে গবেষকরা প্লাস্টিক ভাঙতে ব্যাকটেরিয়া এবং ছত্রাক ব্যবহার শুরু করেন।
মানুষের সাথে সম্পর্ক
পোষাপ্রাণীর খাদ্য হিসাবে
জনপ্রিয় মেলওয়ার্মের মতো জোফোবাস মরিও শূককীট (সাধারণত মহাপোকা নামে পরিচিত) পোষা প্রাণীর যত্নে ব্যাপকভাবে ব্যবহৃত হয় বিশেষত খাদ্য হিসাবে।
মহাপোকাগুলিতে প্রোটিন এবং চর্বি তুলনামূলকভাবে বেশি থাকে, ফলে এগুলি বন্দী সরীসৃপ, উভচর, মাছ এবং পাখিদের জন্য আকর্ষণীয় খাদ্য। এগুলি ১ থেকে ২ সপ্তাহ না খেয়ে বেঁচে থাকতে পারে, এবং তাদের এই ক্ষমতার কারণে বিশ্বজুড়ে পাইকারি পরিমাণে বাণিজ্যিক সুলভতার প্রক্রিয়াটি সহজ। তবে পোষা প্রাণীর মালিকদের এগুলিকে উষ্ণ তাপমাত্রায় রাখার পরামর্শ দেওয়া হয়। কারণ খাবারের কীট থেকে ভিন্ন বলে মহাপোকাগুলি শীতনিদ্রা প্রক্রিয়ায় প্রবেশ করে না। এগুলি নিয়ে কাজ করার সময় হুমকির সম্মূখীন হলে এগুলি কামড়াতেও পারে, যদিও কামড় খুব বেদনাদায়ক নয়।
বর্জ্য নিষ্পত্তিকারক হিসাবে
২০১৬ সালে এটিনিও ডি ম্যানিলা বিশ্ববিদ্যালয়ের ছাত্রদের একটি দল আবিষ্কার করে যে জোফোবাস মোরিওর শূককীট বর্জ্য নিষ্পত্তিতে ব্যবহার করা যেতে পারে। কারণ শূককীটগুলি প্রসারিত পলিস্টাইরিন ফেনা খায় বলে দেখা গেছে। গবেষণা অধ্যয়নটি বৃহত্তর জোফোবাস মরিও শূককীটকে টেনেব্রিও মলিটর শূককীটের সাথে তুলনা করা হয়, যা পূর্বে পলিস্টেরিন অবক্ষয় মোকাবেলা করার জন্য স্ট্যানফোর্ড বিশ্ববিদ্যালয়ের একটি গবেষণায় ব্যবহৃত হয়েছিল
জৈবিক সংগঠন (ইংরেজি: Biological organization) বা জৈবিক সংগঠনের স্তরক্রম বা জীবনের স্তরক্রম (Hierarchy of life) বলতে জীবনকে সংজ্ঞায়িতকারী জটিল জৈবিক কাঠামো ও ব্যবস্থাগুলির একটি স্তরক্রমকে বোঝায়, যাকে জটিল কোনও কিছুকে সরল খণ্ডাংশে বিশ্লিষ্ট করার দৃষ্টিভঙ্গি থেকে নির্মাণ করা হয়েছে।[১] ঐতিহ্যবাহী স্তরক্রমটি পরমাণু থেকে জীবমণ্ডল পর্যন্ত প্রসারিত। এই স্তরক্রমের প্রতিটি স্তরের সংগঠনের মাত্রা তার নিচের স্তরের সাপেক্ষে অধিকতর জটিল। কোনও স্তরের প্রতিটি “বস্তু” বা “সদস্য” মূলত ঠিক তার নিচের স্তরের একাধিক বস্তু বা সদস্যের সমবায়ে গঠিত।[২] এইরূপ সংগঠনের পেছনে যে ভিত্তিসদৃশ মূলনীতিটি কাজ করছে, সেটি হল “উদ্ভব” (Emergence); কোনও নির্দিষ্ট স্তরের সদস্যদের যে ধর্মাবলি ও কাজ পরিলক্ষিত হয়, সেগুলি এর নিচের স্তরগুলিতে অনুপস্থিত থাকে।
জীবনের জৈবিক সংগঠন ও তার স্তরক্রম বৈজ্ঞানিক গবেষণা বহুসংখ্যক ক্ষেত্রের একটি মৌলিক প্রতিজ্ঞা। বিশেষত চিকিৎসাবিজ্ঞানে এর প্রভাব অত্যন্ত গুরুত্বপূর্ণ। যদি জৈবিক সংগঠনের বিভিন্ন মাত্রা সংজ্ঞায়িত না থাকত, তাহলে রোগব্যাধি ও শারীরবিজ্ঞানের গবেষণাতে বিভিন্ন ভৌত ও রাসায়নিক ঘটনার প্রভাব অধ্যয়ন করা দুরূহ এবং সম্ভবত অসম্ভব একটি কাজে পরিণত হত। উদাহরণস্বরূপ, জ্ঞানীয় স্নায়ুবিজ্ঞান ও আচরণীয় স্নায়ুবিজ্ঞানের মতো ক্ষেত্রগুলির হয়ত অস্তিত্বই থাকত না, যদি মানব মস্তিষ্ক কিছু নির্দিষ্ট ধরনের কোষ দিয়ে তৈরি না হত। আবার যদি কোষ পর্যায়ে কোনও পরিবর্তন যদি সমগ্র জীবদেহকে প্রভাবিত না করত, তাহলে ঔষধবিজ্ঞানের মৌলিক ধারণাগুলির কোনও অস্তিত্ব থাকত না। বাস্তুবৈজ্ঞানিক স্তরগুলিতেও একই কথা প্রযোজ্য। যেমন ডিডিটি নামক কীটনাশকের প্রভাব অধোকোষীয় বা কোষনিম্নস্থ স্তরে পরিলক্ষিত হলেও এটি উপরের স্তরগুলিকেও, এমনকি বিভিন্ন বাস্তুতন্ত্রকেও প্রভাবিত করে। তাত্ত্বিকভাবে একটিমাত্র পরমাণুর পরিবর্তন সমগ্র জীবমণ্ডলে পরিবর্তন সৃষ্টি করতে সক্ষম।
স্তরসমূহ
The simplest unit of life is the atom, like oxygen. Two or more atoms is a molecule, like a dioxide. Many small molecules may combine in a chemical reaction to make up a macromolecule, such as a phospholipid. Multiple macromolecules form a cell, like a club cell. A group of cells functioning together as a tissue, for example, Epithelial tissue. Different tissues make up an organ, like a lung. Organs work together to form an organ system, such as the Respiratory System. All of the organ systems make a living organism, like a lion. A group of the same organism living together in an area is a population, such as a pride of lions. Two or more populations interacting with each other form a community, for example, lion and zebra populations interacting with each other. Communities interacting not only with each other but also with the physical environment encompass an ecosystem, such as the Savanna ecosystem. All of the ecosystems make up the biosphere, the area of life on Earth.
নিচে জৈবিক সংগঠনের সরল আদর্শ স্তরক্রমটিকে সর্বনিম্ন স্তর থেকে সর্বোচ্চ স্তর পর্যন্ত উপস্থাপন করা হল:[১]
রাইবোজোম , যাকে ‘প্যালাডে দানা’ ও বলা হয় (আবিষ্কারক জর্জ প্যালাডের[১] নামানুসারে ও দানাদার গঠনের কারণে) এক ধরনের ম্যাক্রোমোলার অঙ্গাণু[২] যা দেহের সকল কোষেই[৩] পাওয়া যায়। এটি মূলত জৈবিক প্রোটিন সংশ্লেষণের[৪] (মেসেঞ্জার আরএনএ বা বার্তাবহ আরএনএ থেকে প্রোটিন তৈরির প্রক্রিয়া) কাজ করে থাকে।
রাইবোজোম পলিপেপটাইড শৃঙ্খল[৫] গঠন করতে বার্তাবহ আরএনএ[৬] এর কোডন[৭] অনুসারে অ্যামিনো অ্যাসিডগুলোর সংযোগ ঘটায়। রাইবোজোম দুটি প্রধান ভাগে বিভক্ত। এরা ছোট ও বড় রাইবোজোমাল সাবইউনিট নামে পরিচিত। প্রতিটি সাবইউনিট এক বা একাধিক রাইবোজোমাল আরএনএ[৮] ও রাইবোজোমাল প্রোটিন[৯] দিয়ে তৈরি।[১০][১১][১২] রাইবোজোম এবং এর সহযোগী অঙ্গাণু সমূহকে ট্রান্সলেশনাল আ্যাপারেটাস বা প্রোটিন তৈরির যন্ত্র নামেও ডাকা হয়।
একনজরে
ডিএনএর[১৩] ক্রম যা একটি প্রোটিনের অ্যামিনো অ্যাসিডের ক্রমকে এনকোড করে তা একটি মেসেঞ্জার আরএনএ শৃঙ্খলে প্রতিলিপি করে। রাইবোসোমগুলি মেসেঞ্জার আরএনএগুলির সাথে আবদ্ধ হয় এবং একটি প্রদত্ত প্রোটিন তৈরি করতে অ্যামিনো অ্যাসিডের সঠিক ক্রম নির্ধারণের জন্য তাদের ক্রমগুলি ব্যবহার করে। অ্যামিনো অ্যাসিডগুলো স্থানান্তরিত আরএনএ (tRNA)[১৪] অণুর মাধ্যমে নির্বাচিত হয় এবং রাইবোসোমে বাহিত হয়, যা রাইবোসোমে প্রবেশ করে এবং অ্যান্টি-কোডন[১৫] স্টেম লুপের মাধ্যমে মেসেঞ্জার আরএনএ শৃঙ্খলের সাথে আবদ্ধ হয়। মেসেঞ্জার আরএনএ-তে প্রতিটি কোডিং ট্রিপলেট (কোডন) এর জন্য, একটি অনন্য ট্রান্সফার আরএনএ রয়েছে যার অবশ্যই সঠিক অ্যান্টি-কোডন মিল থাকতে হবে এবং ক্রমবর্ধমান পলিপেপটাইড চেইনে অন্তর্ভুক্ত করার জন্য সঠিক অ্যামিনো অ্যাসিড বহন করে। প্রোটিন উত্পাদিত হলে, এটি একটি কার্যকরী ত্রিমাত্রিক গঠন তৈরি করতে ভাঁজ[১৬] করতে পারে। একটি রাইবোজোম আরএনএ[১৭] ও অনেকগুলো প্রোটিনের সমন্বয়ে গঠিত এবং এ দুটি একত্রে রাইবোনিউক্লিওপ্রোটিন যৌগ[১৮] তৈরি করে। প্রতিটি রাইবোজোম ছোট ৩০এস এবং বড় ৫০এস সাবইউনিট এ বিভক্ত। উভয়েই একে অপরের উপর নির্ভরশীল।
৩০এস-এর প্রধানত একটি ডিকোডিং ফাংশন রয়েছে এবং এটি এমআরএনএ এর সাথেও আবদ্ধ।
৫০এস-এর প্রধানত একটি অনুঘটক ফাংশন রয়েছে এবং এটি অ্যামিনোঅ্যাসিলেটেড টিআরএনএগুলির সাথেও আবদ্ধ।
তাদের বিল্ডিং ব্লক থেকে প্রোটিনগুলির সংশ্লেষণ চারটি পর্যায়ে সঞ্চালিত হয়: সূচনা, প্রসারণ, সমাপ্তি এবং পুনর্ব্যবহারযোগ্য। সমস্ত এমআরএনএ অণুতে স্টার্ট কোডনের ক্রম AUG আছে। স্টপ কোডন হল UAA, UAG, বা UGA এর মধ্যে একটি; যেহেতু এই কোডনগুলিকে চিনতে পারে এমন কোনও টিআরএনএ অণু নেই, তাই রাইবোসোম মনে করে যে সংশ্লেষণ সম্পূর্ণ হয়েছে।[১৯] যখন একটি রাইবোসোম একটি এমআরএনএ অণু পড়া শেষ করে, তখন দুটি উপইউনিট আলাদা হয়ে যায় এবং সাধারণত ভেঙে যায় তবে পুনরায় ব্যবহার করা যেতে পারে। রাইবোসোমগুলি হল রাইবোজাইম, কারণ অনুঘটক পেপটাইডিল ট্রান্সফারেজের কার্যকলাপ যা অ্যামিনো অ্যাসিডকে একত্রে সংযুক্ত করে তা রাইবোসোমাল আরএনএ দ্বারা সঞ্চালিত হয়।[২০]
রাইবোসোমগুলি প্রায়শই অন্তঃকোষীয় ঝিল্লির সাথে যুক্ত থাকে যা অমসৃণ এন্ডোপ্লাজমিক জালিকা তৈরি করে।
ব্যাকটেরিয়া, আর্কিয়া এবং তিন ডোমেইন সিস্টেমের প্রকৃত কোষের রাইবোজোমের মধ্যে লক্ষণীয় মিল দেখা যায় যা একটি সাধারণ উৎপত্তির প্রমাণ। এদের আকার, আকৃতি, ক্রম, গঠন এবং প্রোটিন ও আরএনএ এর অনুপাত ভিন্ন। গঠনের এই পার্থক্য কিছু ব্যাকটেরিয়া প্রতিরোধীকে (অ্যান্টিবায়োটিক) ব্যাকটেরিয়ার রাইবোজোমকে প্রোটিন সংশ্লেষণে বাধা দেয়ার মাধ্যমে ব্যাকটেরিয়া মারতে সাহায্য করে। যার দরুন মানুষের রাইবোজোম অক্ষুণ্ন থাকে। সকল প্রজাতিতে একের অধিক রাইবোজোম একই সময়ে একটি একক বার্তাবহ আরএনএ এর শিকল বরাবর অগ্রসর হয়। প্রতিটি “রিডিং” একটি নির্দিষ্ট অনুক্রম এবং এরা প্রোটিন অণু উৎপাদন করে।
প্রকৃত কোষের মাইটোকন্ড্রিয়ায় অবস্থিত রাইবোজোম কার্যগতভাবে ব্যাকটেরিয়ার কোষে অবস্থিত রাইবোজোমের অনেক বৈশিষ্ট্যের সাথে মিল দেখায় যা মাইটোকন্ড্রিয়ার বিবর্তনিক উৎপত্তিকেই নির্দেশ করে।[২১][২২]
আবিষ্কার
১৯৫০-এর দশকের মাঝামাঝি সময়ে রোমানিয়ান-আমেরিকান কোষ জীববিজ্ঞানী জর্জ এমিল প্যালাডে, একটি ইলেক্ট্রন অণুবীক্ষণ যন্ত্র ব্যবহার করে ঘন কণা বা দানা হিসেবে রাইবোসোমগুলি প্রথম দেখতে পান।[২৩] “রাইবোসোম” শব্দটি ১৯৫৮ সালের শেষের দিকে বিজ্ঞানী হাগুয়েনাউ প্রস্তাব করেছিলেন:
সিম্পোজিয়াম চলাকালীন একটি অসুবিধা স্পষ্ট হয়ে ওঠে। কিছু অংশগ্রহণকারীদের কাছে, “মাইক্রোসোম” বলতে অন্যান্য প্রোটিন এবং লিপিড উপাদান দ্বারা দূষিত মাইক্রোজোম ভগ্নাংশের রাইবোনিউক্লিওপ্রোটিন কণাকে বোঝায়; অন্যদের কাছে, মাইক্রোসোমগুলি কণা দ্বারা দূষিত প্রোটিন এবং লিপিড নিয়ে গঠিত। শব্দগুচ্ছ “মাইক্রোসোমাল কণা” পর্যাপ্ত বলে মনে হয় না এবং “মাইক্রোসোম ভগ্নাংশের রাইবোনিউক্লিওপ্রোটিন কণা” খুবই বিশ্রী। বৈঠকের সময়, “রাইবোসোম” শব্দটি প্রস্তাবিত হয়েছিল, যেটি খুব সুন্দর নাম এবং শুনতেও ভালো শোনায়। বর্তমান বিভ্রান্তি দূর হবে যদি ৩৫ থেকে ১০০এস পর্যন্ত আকারে রাইবোনিউক্লিওপ্রোটিন কণা নির্ধারণের জন্য “রাইবোসোম” নামটি গ্রহণ করা হয়।[২৪]
অ্যালবার্ট ক্লদ, ক্রিশ্চিয়ান দ্য দুবে এবং জর্জ এমিল প্যালাডে ১৯৭৪ সালে রাইবোসোম আবিষ্কারের জন্য যৌথভাবে চিকিৎসা বিজ্ঞানে নোবেল পুরস্কার লাভ করেন।[২৫] রাইবোসোমের বিশদ গঠন ও প্রক্রিয়া নির্ধারণের জন্য ২০০৯ সালে রসায়নে নোবেল পুরস্কার দেওয়া হয় ভেঙ্কটরামন রামকৃষ্ণান, টমাস এ. স্টিটজ এবং অ্যাডা ই. ইয়োনাথকে।[২৬]
গঠন
রাইবোসোম একটি জটিল কোষীয় অঙ্গাণু। এটি মূলত রাইবোসোমাল আরএনএ (rRNA) এবং কয়েক ডজন স্বতন্ত্র প্রোটিন দ্বারা গঠিত (সঠিক সংখ্যা প্রজাতির মধ্যে সামান্য পরিবর্তিত হয়)। রাইবোসোমাল প্রোটিন এবং আরআরএনএগুলি বিভিন্ন আকারের দুটি স্বতন্ত্র রাইবোসোমাল টুকরোতে বিন্যস্ত থাকে, যা সাধারণত রাইবোসোমের বড় এবং ছোট সাবইউনিট হিসাবে পরিচিত। রাইবোসোম দুটি সাবইউনিট নিয়ে গঠিত যা একসাথে সংযুক্ত থাকে এবং প্রোটিন সংশ্লেষণের সময় এমআরএনএ -কে একটি পলিপেপটাইড চেইনে রূপান্তর করতে কাজ করে। যেহেতু তারা অ-সমান আকারের দুটি সাবইউনিট থেকে গঠিত, তাই তারা ব্যাসের তুলনায় অক্ষে কিছুটা লম্বা।
আদিকোষী রাইবোসোম
আদিকোষী রাইবোসোমগুলির ব্যাস প্রায় ২০ ন্যানোমিটার (২০০ Å) এবং ৬৫% রাইবোসোমাল আরএনএ এবং ৩৫% রাইবোসোমাল প্রোটিন[২৭] দ্বারা গঠিত। প্রকৃত কোষের রাইবোসোমগুলির ব্যাস ২৫ থেকে ৩০ ন্যানোমিটার(২৫০-৩০০ Å) এর মধ্যে এবং এতে আর আরএনএ ও প্রোটিনের অনুপাত ১ এর কাছাকাছি।[২৮] ক্রিস্টালোগ্রাফি[২৯] করে দেখা গিয়েছে যে পলিপেপটাইড সংশ্লেষণের জন্য প্রতিক্রিয়া স্থানের কাছাকাছি কোনও রাইবোসোমাল প্রোটিন নেই। এ থেকে বোঝা যায় রাইবোসোমের প্রোটিন উপাদানগুলি পেপটাইড বন্ধন গঠনের অনুঘটকগুলিতে সরাসরি অংশগ্রহণ করে না, বরং এই প্রোটিনগুলি একটি ভারা হিসাবে কাজ করে যা প্রোটিন সংশ্লেষণ করার জন্য আর আরএনএ এর ক্ষমতা বাড়ায়। (দেখুন: রাইবোজাইম)।
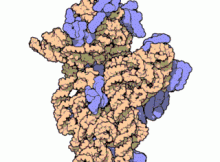
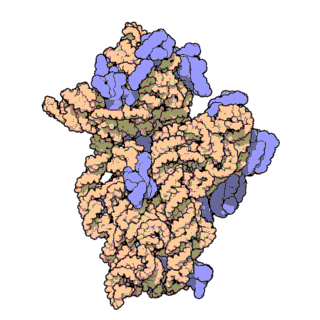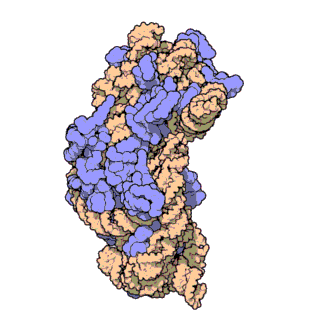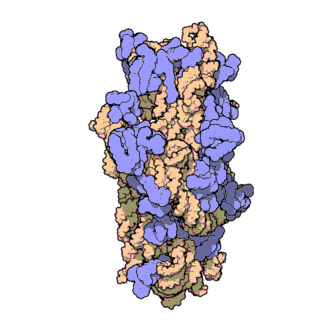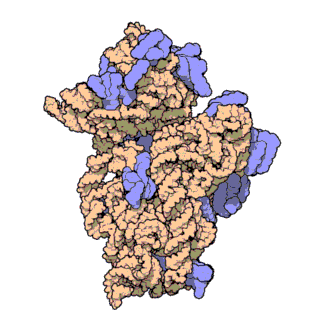
চিত্র ৩: থার্মাস থার্মোফিলাস থেকে ৩০এস সাবইউনিটের আণবিক গঠন। প্রোটিনগুলি নীল রঙে এবং একক আরএনএ শৃঙ্খল বাদামী রঙে দেখানো হয়েছে।
আদিকোষী এবং প্রকৃত কোষী রাইবোসোমাল সাবইউনিটগুলি দেখতে বেশ একই রকম।[৩০]
রাইবোসোমাল সাবইউনিট এবং আর- আরএনএ এর খণ্ডগুলি বর্ণনা করতে ব্যবহৃত পরিমাপের একক হল ভেদবার্গ (Svedberg) একক। এর মাধ্যমে আকারের পরিবর্তে কেন্দ্রীভূতকরণে অবক্ষেপণের হারের একটি পরিমাপ করা হয়। উদাহরণস্বরূপ, ব্যাকটেরিয়ার ৭০এস রাইবোসোমগুলি ৫০এস এবং ৩০এস সাবইউনিট দিয়ে তৈরি।
আদিকোষে ৭০এস রাইবোসোম আছে, প্রতিটি একটি ছোট (৩০এস) এবং একটি বড় (৫০এস) সাবইউনিট নিয়ে গঠিত।উদাহরণস্বরূপ, ই-কোলাই , একটি ১৬এস আরএনএ সাবইউনিট (১৫৪০ নিউক্লিওটাইড সমন্বিত) যা ২১টি প্রোটিনের সাথে আবদ্ধ। বৃহৎ সাবইউনিট একটি ৫এস আরএনএ সাবইউনিট (১২০ নিউক্লিওটাইড), একটি ২৩এস আরএনএ উপইউনিট (২৯০০ নিউক্লিওটাইড) এবং ৩১টি প্রোটিন দ্বারা গঠিত।[৩০]
রাইবোসোম
সাবইউনিট
আর আরএনএ
আর প্রোটিন
৭০এস
৫০এস
২৩এস (২৯০৪ নিউক্লিওটাইড)
৩১
৩০এস
৫এস (১২০নিউক্লিওটাইড)
২১
১৬এস (১৫৪২নিউক্লিওটাইড)
ই কোলাই রাইবোসোমে টি আরএনএ বাইন্ডিং সাইটগুলির জন্য অ্যাফিনিটি লেবেল এ এবং পি সাইট প্রোটিন সনাক্ত করার অনুমতি দেয় ।সম্ভবত পেপটাইডাইলট্রান্সফেরেজ কার্যকলাপের[৩১] সাথে যুক্ত;লেবেলযুক্ত প্রোটিনগুলি হল এল২৭, এল১৪, এল১৫, এল১৬, এল২। এল২৭ ডোনার সাইটে অবস্থিত। যেমনটি ই. কোলাটজ এবং এপি চের্নিলোফস্কি[৩২][৩৩] -এর গবেষণায় প্রমাণিত হয়েছে যে এস১ ও এস২১ প্রোটিন, ১৬এস রাইবোসোমাল আরএনএ-এর ৩′-শেষের সাথে যুক্ত অনুবাদের সূচনাতে জড়িত।[৩৪]
আর্কিয়াল রাইবোসোম
আর্কিয়াল রাইবোসোমগুলি ব্যাকটেরিয়াগুলির একই সাধারণ মাত্রা ভাগ করে, একটি ৭০S রাইবোসোম যা একটি ৫০এস বড় সাবইউনিট, একটি ৩০এস ছোট সাবইউনিট থেকে গঠিত এবং তিনটিআর আরএনএ শৃঙ্খল রয়েছে। যাইহোক, ক্রম স্তরে, তারা ব্যাকটেরিয়ার তুলনায় প্রকৃত কোষগুলির অনেক কাছাকাছি। ব্যাকটেরিয়ার সাথে তুলনা করা আর্কিয়ার প্রতিটি অতিরিক্ত রাইবোসোমাল প্রোটিনের একটি প্রকৃত কোষী প্রতিরূপ রয়েছে, যখন আর্কিয়া এবং ব্যাকটেরিয়ার মধ্যে এই ধরনের কোন সম্পর্ক প্রযোজ্য নয়।[৩৫][৩৬][৩৭]
প্রকৃত কোষী রাইবোসোম
প্রকৃত কোষের সাইটোসোলে ৮০এস রাইবোসোম থাকে, প্রতিটিতে একটি ছোট (৪০এস) এবং বড় (৬০এপ) সাবইউনিট থাকে। তাদের ৪০এস সাবইউনিটে একটি ১৮এস আরএনএ (১৯০০ নিউক্লিওটাইড) এবং ৩৩টি প্রোটিন রয়েছে।[৩৮] বৃহৎ সাবইউনিট একটি ৫এস আরএনএ (১২০টি নিউক্লিওটাইড), ২৮এস আরএনএ (৪৭০০টি নিউক্লিওটাইড), একটি ৫.৮এস আরএনএ (১৬০টি নিউক্লিওটাইড) সাবইউনিট এবং ৪৬টি প্রোটিন দ্বারা গঠিত।[৩০][৩৮][৩৯]
রাইবোসোম
সাবইউনিট
আর আরএনএ
আর প্রোটিন
৮০এস
৬০এস
২৮এস (৪৭১৮নিউক্লিওটাইড)
৪৯
৫.৮এস (১৬০নিউক্লিওটাইড)
৫এস (১২০ নিউক্লিওটাইড)
৩৩
৪০এস
১৮এস (১৮৭৪নিউক্লিওটাইড)
১৯৭৭ সালে, চের্নিলোফস্কি গবেষণা প্রকাশ করেছিলেন যা ইঁদুরের যকৃতের রাইবোসোমগুলিতে টিআরএনএ-বাইন্ডিং সাইটগুলি সনাক্ত করতে অ্যাফিনিটি লেবেলিং ব্যবহার করেছিল। এল৩২/৩৩, এল৩৬, এল২১, এল২৩, এল২৮/২৯ এবং এল১৩ সহ বেশ কিছু প্রোটিন পেপটাইডিল ট্রান্সফারেজ কেন্দ্রে[৪০] বা তার কাছাকাছি হিসাবে জড়িত ছিল।
প্লাস্টোরিবোসোম এবং মাইটোরিবোসোম
প্রকৃত কোষে, রাইবোসোমগুলি মাইটোকন্ড্রিয়াতে (কখনও কখনও মাইটোরিবোসোম বলা হয়) এবং ক্লোরোপ্লাস্টের মতো প্লাস্টিডে (যাকে প্লাস্টোরিবোসোমও বলা হয়) উপস্থিত থাকে। তারা একটি ৭০S কণাতে প্রোটিনের সাথে একসাথে আবদ্ধ বড় এবং ছোট সাবইউনিট নিয়ে গঠিত। এই রাইবোসোমগুলি ব্যাকটেরিয়াগুলির অনুরূপ এবং এই অঙ্গাণুগুলি মিথোজীবী ব্যাকটেরিয়া[৩০] হিসাবে উদ্ভূত হয়েছে বলে মনে করা হয় । মাইটোক্রন্ড্রিয়ায় রাইবোসোমাল RNA-এর অনেকগুলি টুকরো ছোট করা হয়, এবং ৫S rRNA-এর ক্ষেত্রে, প্রাণী ও ছত্রাকের[৪১] অন্যান্য গঠন দ্বারা প্রতিস্থাপিত হয়। বিশেষ করে, লেইশম্যানিয়া ট্যারেন্টোলে মাইটোকন্ড্রিয়াল rRNA এর একটি সংক্ষিপ্ত সেট রয়েছে।[৪২] বিপরীতে, উদ্ভিদের মাইটোরিবোসোম আছে b ব্যাকটেরিয়ার তুলনায় অন্যান্য বর্ধিত rRNA এবং অতিরিক্ত প্রোটিন, বিশেষ করে, অনেক পেন্টাট্রিকোপেটাইড পুনরাবৃত্তি প্রোটিন।[৪৩]
ক্রিপ্টোমোনাড এবং ক্লোররাচনিওফাইট শৈবালের মধ্যে একটি নিউক্লিওমর্ফ থাকতে পারে যা একটি ভেস্টিজিয়াল প্রকৃত কোষী নিউক্লিয়াসের[৪৪] মতো। প্রকৃত কোষী ৮০S রাইবোসোম নিউক্লিওমর্ফ ধারণকারী বগিতে উপস্থিত থাকতে পারে।[৪৫]
পার্থক্য ব্যবহার করা
ব্যাকটেরিয়া এবং প্রকৃত কোষী রাইবোসোমের মধ্যে পার্থক্যগুলি ঔষধ তৈরিকারী রসায়নবিদদের দ্বারা অ্যান্টিবায়োটিক তৈরির জন্য ব্যবহার করা হয় যা সংক্রামিত ব্যক্তির কোষের ক্ষতি না করেই ব্যাকটেরিয়া সংক্রমণকে ধ্বংস করতে পারে। তাদের গঠনগত পার্থক্যের কারণে, ব্যাকটেরিয়া ৭০S রাইবোসোমগুলি এই অ্যান্টিবায়োটিকের জন্য ঝুঁকিপূর্ণ যেখানে ইউক্যারিওটিক ৮০S রাইবোসোমগুলি নয়।[৪৬] যদিও মাইটোকন্ড্রিয়াতে ব্যাকটেরিয়ার অনুরূপ রাইবোসোম থাকে, মাইটোকন্ড্রিয়া এই অ্যান্টিবায়োটিকের দ্বারা প্রভাবিত হয় না কারণ তারা একটা দ্বিস্তরী ঝিল্লি দ্বারা বেষ্টিত থাকে যা এই অ্যান্টিবায়োটিকগুলিকে সহজেই অঙ্গাণুর মধ্যে প্রবেশ করতে দেয় না। একটি উল্লেখযোগ্য উদাহরণ, তবে, অ্যান্টিনিওপ্লাস্টিক অ্যান্টিবায়োটিক ক্লোরামফেনিকল অন্তর্ভুক্ত করে, যা সফলভাবে ব্যাকটেরিয়া ৫০এস এবং প্রকৃত কোষী মাইটোকন্ড্রিয়াল ৫০এস রাইবোসোমকে বাধা দেয়। মাইটোকন্ড্রিয়ার একই ব্যাপারটি ক্লোরোপ্লাস্টের ক্ষেত্রে ঘটে না, যেখানে রাইবোসোমাল প্রোটিনে অ্যান্টিবায়োটিক প্রতিরোধের একটি বৈশিষ্ট্য যা জিন প্রকৌশলে চিহ্নিতকারী হিসাবে ব্যবহৃত হয়।[৪৭]
সাধারণ বৈশিষ্ট্য
বিভিন্ন রাইবোসোম একটি মূল কাঠামো ভাগ করে, যা আকারে বড় পার্থক্য থাকা সত্ত্বেও বেশ কিছু মিল আছে। আরএনএ-এর বেশিরভাগ অংশই বিভিন্ন তিন স্তরের কাঠামোগত মোটিফগুলিতে অত্যন্ত সংগঠিত, উদাহরণস্বরূপ সিউডোকনট যা সমাক্ষীয় স্ট্যাকিং প্রদর্শন করে। বৃহত্তর রাইবোসোমের অতিরিক্ত আরএনএ বেশ কিছু দীর্ঘ একটানা সন্নিবেশে থাকে,[৪৮] যাতে তারা মূল কাঠামোর বাইরে লুপ তৈরি করে তা ব্যাহত বা পরিবর্তন না করে। রাইবোসোমের সমস্ত অনুঘটক কার্যকলাপ RNA দ্বারা সঞ্চালিত হয়; প্রোটিন পৃষ্ঠের উপর থাকে এবং গঠন স্থিতিশীল বলে মনে হয়।[৩০]
উচ্চ-রেজোলিউশন কাঠামো
পারমাণবিক রেজোলিউশনে রাইবোসোমের গঠন প্রদানকারী প্রথম কাগজপত্রগুলি প্রায় ২০০০ সালের শেষের দিকে প্রায় একই সাথে প্রকাশিত হয়েছিল। ৫০এস (বড় আদিকোষী) সাবইউনিটটি Haloarculamarismortui এবং ব্যাকটেরিয়া Deinococcus radiodurans এর গঠন থেকে নির্ধারণ করা হয়েছিল। ৩০এস সাবইউনিট থার্মাস থার্মোফিলাস থেকে নির্ধারিত হয়েছিল। এই কাঠামোগত গবেষণাটি ২০০৯ সালে রসায়নে নোবেল পুরস্কারে ভূষিত হয়েছিল। মে ২০০১-এ এই স্থানাঙ্কগুলি ৫.৫ Å রেজোলিউশনে সমগ্র টি থার্মোফিলাস ৭০S কণা পুনর্গঠনের জন্য ব্যবহার করা হয়েছিল। ২০০৫ সালের নভেম্বরে ইশারশিয়া কোলাই এর ৭০S রাইবোসোমের গঠন নিয়ে দুটি গবেষণাপত্র প্রকাশিত হয়েছিল। এক্স-রে ক্রিস্টালোগ্রাফি ব্যবহার করে একটি খালি রাইবোসোমের গঠন ৩.৫ Å রেজোলিউশনে নির্ধারণ করা হয়েছিল। [৪৯] তারপরে, দুই সপ্তাহ পরে, ক্রায়ো-ইলেক্ট্রন মাইক্রোস্কোপির উপর ভিত্তি করে একটি কাঠামো প্রকাশিত হয়েছিল, যা প্রোটিন-পরিবাহী চ্যানেলে একটি নতুন সংশ্লেষিত প্রোটিন স্ট্র্যান্ড পাস করার কাজে ১১-১৫ Å রেজোলিউশনে রাইবোসোমকে চিত্রিত করে। টিআরএনএ এবং এমআরএনএ অণুগুলির সাথে জটিল রাইবোসোমের প্রথম পারমাণবিক কাঠামো দুটি স্বাধীনভাবে ২.৮ Å এবং ৩.৭ Å এ এক্স-রে ক্রিস্টালোগ্রাফি ব্যবহার করে সমাধান করা হয়েছিল। এই কাঠামোগুলি একজনকে থার্মাস থার্মোফিলাস রাইবোসোমের সাথে mRNA এবং ক্লাসিক্যাল রাইবোসোমাল স্থানগুলিতে আবদ্ধ tRNA এর সাথে মিথস্ক্রিয়াগুলির বিশদ বিবরণ দেখতে দেয়। শাইন-ডালগার্নো সিকোয়েন্স ধারণকারী দীর্ঘ mRNA-এর সাথে রাইবোসোমের মিথস্ক্রিয়া ৪.৫-৫.৫ Å রেজোলিউশনে এর পরেই কল্পনা করা হয়েছিল।[৫০] ২০১১ সালে, এক ধরনের ঈস্ট Saccharomyces cerevisiae থেকে প্রকৃত কোষীদের ৮০S রাইবোসোমের প্রথম সম্পূর্ণ পারমাণবিক কাঠামো ক্রিস্টালোগ্রাফির মাধ্যমে পাওয়া গিয়েছিল। মডেলটি প্রকৃত কোষীদের -নির্দিষ্ট উপাদানগুলির গঠন এবং সর্বজনীনভাবে সংরক্ষিত মূলের সাথে তাদের মিথস্ক্রিয়া প্রকাশ করে। একই সময়ে, টেট্রাহাইমেনা থার্মোফিলায় একটি প্রকৃত কোষীদের ৪০S রাইবোসোমাল কাঠামোর সম্পূর্ণ মডেল প্রকাশিত হয়েছিল এবং ৪০S সাবইউনিটের গঠন বর্ণনা করা হয়েছিল । সেইসাথে সংশ্লেষণ শুরুর সময় eIF১ এর সাথে ৪০S সাবইউনিটের মিথস্ক্রিয়া সম্পর্কে অনেক কিছু বলা হয়েছিল।একইভাবে, প্রকৃত কোষী ৬০S সাবইউনিট গঠনটিও টেট্রাহাইমেনা থার্মোফিলা থেকে eIF৬[৫১] এর সাথে কমপ্লেক্সে নির্ধারিত হয়েছিল।
কার্যাবলী
রাইবোসোম হল ক্ষুদ্র ক্ষুদ্র কণা যার মধ্যে আরএনএ এবং সংশ্লিষ্ট প্রোটিন রয়েছে যা প্রোটিন সংশ্লেষণের জন্য কাজ করে। প্রোটিন কোষস্থ বিভিন্ন কার্যাবলী যেমন ক্ষতি মেরামত বা রাসায়নিক প্রক্রিয়া নির্দেশ করার জন্য প্রয়োজন হয়। রাইবোসোমগুলি সাইটোপ্লাজমের মধ্যে ভাসমান বা এন্ডোপ্লাজমিক রেটিকুলামের সাথে সংযুক্ত পাওয়া যায়। তাদের প্রধান কাজ হল জেনেটিক কোডকে অ্যামিনো অ্যাসিড সিকোয়েন্সে রূপান্তর করা এবং অ্যামিনো অ্যাসিডের মনোমার থেকে প্রোটিন পলিমার তৈরি করা।
রাইবোসোম দুটি অত্যন্ত গুরুত্বপূর্ণ জৈবিক প্রক্রিয়ায় অনুঘটক হিসেবে কাজ করে যার নাম পেপটাইডিল স্থানান্তর এবং পেপটাইডিল পানি বিশ্লেষণ। “PT কেন্দ্র প্রোটিন প্রসারণের সময় প্রোটিন বন্ধন তৈরির জন্য দায়ী”।[৫২]
সংক্ষেপে, রাইবোসোমের দুটি প্রধান কাজ রয়েছে: বার্তা পাঠোদ্ধার করা এবং পেপটাইড বন্ধন গঠন করা। এই দুটি ফাংশন রাইবোসোমাল সাবইউনিটে থাকে। প্রতিটি সাবইউনিট এক বা একাধিক রাইবোসোমাল আরএনএ এবং অনেকগুলি আর -প্রোটিন দিয়ে তৈরি। ছোট সাবইউনিট (ব্যাকটেরিয়া এবং আর্কিয়াতে ৩০এস, প্রকৃত কোষে ৪০S) ডিকোডিং ফাংশন রয়েছে, যেখানে বড় সাবইউনিট (ব্যাকটেরিয়া এবং আর্কিয়াতে ৫০এস, প্রকৃত কোষে ৬০S) পেপটাইড বন্ধন গঠনকে প্রভাবিত করে, যাকে পেপটাইডিল-ট্রান্সফারেজ কার্যকলাপ বলা হয়। ব্যাকটেরিয়া (এবং আর্কিয়াল) ছোট সাবইউনিটে ১৬S আর আরএনএ এবং ২১ আর-প্রোটিন (Escherichia coli) থাকে, যেখানে প্রকৃত কোষের ছোট সাবইউনিটে ১৮S আর আরএনএ এবং ৩২ আর-প্রোটিন থাকে (স্যাকারোমাইসেস সেরিভিসিয়া; যদিও প্রজাতির মধ্যে সংখ্যার ভিন্নতা দেখা যায়)। ব্যাকটেরিয়াল বৃহৎ সাবইউনিটে ৫S এবং ২৩S আর আরএনএ এবং ৩৪টি আর-প্রোটিন (E. coli), প্রকৃত কোষের বৃহৎ সাবইউনিটে রয়েছে ৫S, ৫.৮S এবং ২৫S/২৮S আর আরএনএ এবং ৪৬টি আর-প্রোটিন (উদাহরণ S. cerevisiae; এক্ষেত্রেও সঠিক সংখ্যা প্রজাতির মধ্যে পরিবর্তিত হয়)।[৫৩]
সংশ্লেষণ
রাইবোসোম হল প্রোটিন জৈব সংশ্লেষণের কারখানা, যেখানে এমআরএনএকে প্রোটিনে রূপান্তরিত করা হয়। এম আরএনএ-তে কোডনগুলির একটি সিরিজ রয়েছে যা প্রোটিন তৈরি করার জন্য রাইবোসোম দ্বারা ডিকোড করা হয়। একটি টেমপ্লেট হিসাবে এম আরএনএ কে ব্যবহার করে, রাইবোসোম এম আরএনএ এর প্রতিটি কোডন (৩ নিউক্লিওটাইড) অতিক্রম করে, এটিকে একটি অ্যামিনোঅ্যাসিল-টি আরএনএ দ্বারা প্রদত্ত উপযুক্ত অ্যামিনো অ্যাসিডের সাথে যুক্ত করে। অ্যামিনোঅ্যাসাইল টি আরএনএ এর এক প্রান্তে একটি পরিপূরক অ্যান্টিকোডন এবং অন্য প্রান্তে উপযুক্ত অ্যামিনো অ্যাসিড থাকে। উপযুক্ তটি আরএনএ -এর দ্রুত এবং সঠিক স্বীকৃতির জন্য, রাইবোসোম বৃহৎ গঠনমূলক পরিবর্তন (কনফরমেশনাল প্রুফরিডিং) ব্যবহার করে।[৫৪]
ছোট রাইবোসোমাল সাবইউনিট, সাধারণত প্রথম অ্যামিনো অ্যাসিড মেথিওনিন ধারণকারী একটি অ্যামিনোঅ্যাসিল-টিআরএনএ-তে আবদ্ধ, এমআরএনএ-তে একটি AUG কোডনের সাথে আবদ্ধ হয় এবং বড় রাইবোসোমাল সাবইউনিট নিয়োগ করে। রাইবোসোমে তিনটি আরএনএ বাইন্ডিং সাইট রয়েছে, মনোনীত A, P এবং E। A-সাইটটি একটি অ্যামিনো অ্যাসিল-টি আরএনএ বা পরিসমাপ্তি রিলিজ ফ্যাক্টরকে আবদ্ধ করে; পলি-পেপটাইড চেইন); এবং ই-সাইট একটি মুক্ত টিআরএনএ আবদ্ধ করে। [৫৫] প্রোটিন সংশ্লেষণ এম আরএনএ এর ৫’ প্রান্তের কাছে একটি স্টার্ট কোডন AUG থেকে শুরু হয়। এম আরএনএ প্রথমে রাইবোসোমের P সাইটে আবদ্ধ হয়। রাইবোসোম আদিকোষে এমআরএনএর শাইন-ডালগারনো ক্রম এবং প্রকৃত কোষে কোজাক বক্স ব্যবহার করে স্টার্ট কোডনকে চিনতে পারে।
যদিও পেপটাইড বন্ধনের অনুঘটক একটি প্রোটন শাটল পদ্ধতিতে আরএনএ-এর P-সাইট অ্যাডিনোসিনের C২ হাইড্রক্সিলকে জড়িত করে, প্রোটিন সংশ্লেষণের অন্যান্য ধাপগুলি (যেমন ট্রান্সলোকেশন) প্রোটিন গঠনের পরিবর্তনের কারণে ঘটে। যেহেতু তাদের অনুঘটক কোর আরএনএ দিয়ে তৈরি, তাই রাইবোসোমগুলিকে “রাইবোজাইম” হিসাবে শ্রেণীবদ্ধ করা হয়,[৫৬] এবং ধারণা করা হয় যে তারা আরএনএ জগতের অবশিষ্টাংশ হতে পারে।[৫৭]
কোট্রান্সলেশনাল ভাঁজ
রাইবোসোম সক্রিয়ভাবে প্রোটিন ভাঁজ করায় অংশ নেয়।[৫৮] এইভাবে প্রাপ্ত গঠনগুলি সাধারণত প্রোটিনের রাসায়নিক পুনঃভাঁজের সময় প্রাপ্ত গঠনের সাথে অভিন্ন। তবে, সর্বশেষ উৎপাদিত পদার্থের গমন পথ ভিন্ন হতে পারে ।কিছু ক্ষেত্রে, রাইবোসোম কার্যকরী প্রোটিনের রূপ প্রাপ্তির জন্য অত্যন্ত গুরুত্বপূর্ণ। উদাহরণস্বরূপ, জটিলভাবে গিঁটযুক্ত প্রোটিনগুলির ভাঁজ করার সম্ভাব্য প্রক্রিয়াগুলির মধ্যে একটি রাইবোসোমের উপর নির্ভর করে যা সংযুক্ত লুপের মাধ্যমে চেইনকে ঠেলে দেয়।[৫৯]
সংশ্লেষণকারী-স্বাধীন অ্যামিনো অ্যাসিডের সংযোজন
একটি রাইবোসোমের মান নিয়ন্ত্রণ প্রোটিন Rqc২ উপস্থিতি এম আরএনএ -স্বাধীন প্রোটিন প্রসারণের সাথে যুক্ত। এই প্রসারণটি ক্যাট লেজের রাইবোসোমাল সংযোজনের (Rqc২ দ্বারা আনা tRNA এর মাধ্যমে) ফল: রাইবোসোমগুলি অ্যালানিন এবং থ্রোনিনের এলোমেলো, অনুবাদ-স্বাধীন ক্রম সহ একটি স্থবির প্রোটিনের সি-টার্মিনাসকে প্রসারিত করে।[৬০]
রাইবোসোমের অবস্থান
রাইবোসোমগুলিকে “মুক্ত” বা “ঝিল্লিবদ্ধ” হিসাবে শ্রেণীবদ্ধ করা হয়।
মুক্ত এবং ঝিল্লিবদ্ধ রাইবোসোমগুলির মধ্যে পার্থক্য শুধুমাত্র তাদের স্থানিক বন্টনের মধ্যে ; তাদের গঠন অভিন্ন । রাইবোসোম একটি মুক্ত বা ঝিল্লি-বাউন্ড অবস্থায় বিদ্যমান কিনা তা প্রোটিন সংশ্লেষিত হওয়ার উপর একটি ER- টার্গেটিং সিগন্যাল সিকোয়েন্সের উপস্থিতির উপর নির্ভর করে, তাই একটি পৃথক রাইবোসোম একটি প্রোটিন তৈরি করার সময় ঝিল্লিতে আবদ্ধ হতে পারে, কিন্তু আরেকটা প্রোটিন তৈরি করার সময় এটি কোষরসে মুক্ত অবস্থায় থাকে।
রাইবোসোমগুলিকে কখনও কখনও অঙ্গাণু হিসাবে উল্লেখ করা হয়, তবে অঙ্গাণু শব্দটির ব্যবহার প্রায়শই একটি ফসফোলিপিড ঝিল্লির অন্তর্ভুক্ত সাব-সেলুলার উপাদানগুলিকে বর্ণনা করার জন্য সীমাবদ্ধ থাকে, কিন্তু রাইবোসোমগুলি সেভাবে থাকে না। এই কারণে, রাইবোসোমগুলিকে কখনও কখনও “ঝিল্লি বিহীন অঙ্গাণু ” হিসাবে বর্ণনা করা যেতে পারে।
মুক্ত রাইবোসোম
মুক্ত রাইবোসোমগুলি কোষরসের যে কোনও জায়গায় ঘুরতে পারে, তবে কোষের নিউক্লিয়াস এবং অন্যান্য অঙ্গাণুর বাইরে থাকে। মুক্ত রাইবোসোম থেকে গঠিত প্রোটিন কোর্সে মুক্তি পায় এবং কোষের ভেতরেই ব্যবহৃত হয়। যেহেতু সাইটোসল গ্লুটাথিয়নের উচ্চ ঘনত্ব ধারণ করে সেহেতু এটি একটি হ্রাসকারী পরিবেশ, ডাইসালফাইড বন্ধন ধারণকারী প্রোটিন, যা অক্সিডাইজড সিস্টাইন অবশিষ্টাংশ থেকে গঠিত হয়, এর মধ্যে উৎপাদিত হতে পারে না।
ঝিল্লিবদ্ধ রাইবোসোম
যখন একটি রাইবোসোম কিছু অঙ্গাণুতে প্রয়োজনীয় প্রোটিন সংশ্লেষণ করতে শুরু করে, তখন এই প্রোটিন তৈরি করা রাইবোসোম “ঝিল্লিবদ্ধ” হয়ে যেতে পারে। প্রকৃত কোষে এটি এন্ডোপ্লাজমিক জালিকার (ER) একটি অঞ্চলে ঘটে যাকে “অমসৃণ এন্ডোপ্লাজমিক জালিকা” বলা হয়। নতুন উৎপাদিত পলিপেপটাইড চেইনগুলি রাইবোসোমের মাধ্যমে আন্ডারটেকিং ভেক্টরিয়াল সংশ্লেষণ দ্বারা সরাসরি এন্ডোপ্লাজমিক জালিকাতে ঢোকানো হয় এবং তারপর নিঃসৃত পথের মাধ্যমে তাদের গন্তব্যে স্থানান্তরিত হয়। আবদ্ধ রাইবোসোমগুলি সাধারণত প্রোটিন তৈরি করে যা কোষ ঝিল্লির মধ্যে ব্যবহৃত হয় বা এক্সোসাইটোসিসের মাধ্যমে কোষ থেকে বেরিয়ে যায়।[৩০]
বায়োজেনেসিস
ব্যাকটেরিয়া কোষে, একাধিক রাইবোসোমাল জিন অপেরনের ট্রান্সক্রিপশনের মাধ্যমে রাইবোসোমগুলি সাইটোপ্লাজমে সংশ্লেষিত হয়। প্রকৃত কোষে প্রক্রিয়াটি কোষের সাইটোপ্লাজম এবং নিউক্লিওলাস (নিউক্লিয়াসের মধ্যে একটি অঞ্চল)উভয় স্থানেই ঘটে।এ সমাবেশ প্রক্রিয়ায় চারটি রাইবোসোমাল আরএনএ এর সংশ্লেষণ এবং প্রক্রিয়াকরণে ২০০ টিরও বেশি প্রোটিনের সমন্বিত ফাংশন জড়িত থাকে, সেইসাথে রাইবোসোমাল প্রোটিনের সাথে সেই আর আরএনএ গুলিকে যুক্ত করে।
উৎপত্তি
রাইবোসোম প্রথম একটি আরএনএ এর জগতে উদ্ভূত হতে পারে। এটি একটি স্ব-প্রতিলিপিকারী কমপ্লেক্স হিসাবে আবির্ভূত হয় যা পরবর্তীতে অ্যামিনো অ্যাসিড উপস্থিত হতে শুরু করলে এর মধ্যে প্রোটিন সংশ্লেষণ করার ক্ষমতা বিকশিত হয়।[৬১] গবেষণায় দেখা গেছে যে শুধুমাত্র আর আরএনএ দ্বারা নির্মিত প্রাচীন রাইবোসোমগুলি পেপটাইড বন্ধন সংশ্লেষণ করার ক্ষমতা তৈরি করতে পারে।[৬২][৬৩] উপরন্তু, প্রমাণগুলি দৃঢ়ভাবে প্রাচীন রাইবোসোমকে স্ব-প্রতিলিপিকারী কমপ্লেক্স হিসাবে নির্দেশ করে[৬৪], যেখানে রাইবোসোমের আর আরএনএ -এর তথ্যগত, কাঠামোগত এবং প্রভাবকীয় উদ্দেশ্য ছিল । কারণ এটি রাইবোসোমাল স্ব-প্রতিলিপির জন্য প্রয়োজনীয় টি আরএনএ এবং প্রোটিনের জন্য কোড করতে সক্ষম। ডিএনএ ছাড়া স্ব-প্রতিলিপিকারী আরএনএ জীবকে রাইবোসাইট (বা রাইবোসেল) বলা হয়।
যেহেতু অ্যামিনো অ্যাসিডগুলি ধীরে ধীরে প্রিবায়োটিক অবস্থার অধীনে আরএনএ বিশ্বে আবির্ভূত হয়, অনুঘটক আরএনএর সাথে তাদের মিথস্ক্রিয়া অনুঘটক আরএনএ অণুর কাজের পরিসর এবং কার্যকারিতা উভয়ই বাড়িয়ে তোলে।[৬৫][৬৬] এভাবে বিবর্তনের জন্য রাইবোসোমের চালিকাশক্তি একটি প্রাচীন স্ব-প্রতিলিপনকারী মেশিন থেকে বর্তমানের একটি প্রোটিন সংশ্লেষণ মূলক যন্ত্রে পরিণত করেছে। নির্বাচনী চাপ রাইবোসোমের স্ব-প্রতিলিপন প্রক্রিয়ায় প্রোটিনগুলিকে যেমন অন্তর্ভুক্ত করে, তেমনি তার প্রতিলিপনের সক্ষমতা বাড়িয়ে দেয়।[৬৭][৬৮]
বিভিন্ন রাইবোসোম
রাইবোসোমগুলি গঠনগত দিক থেকে প্রজাতির মধ্যে এমনকি একই কোষের মধ্যেও ভিন্ন ভিন্ন। যেমনটি একই প্রকৃত কোষের মধ্যকার সাইটোপ্লাজমিক এবং মাইটোকন্ড্রিয়া রাইবোসোমের ভেতর পার্থক্য দেখা যায় । কিছু গবেষক মনে করেন স্তন্যপায়ী প্রাণীদের মধ্যে রাইবোসোমাল প্রোটিনের গঠনের ভিন্নতা জিন নিয়ন্ত্রণের জন্য গুরুত্বপূর্ণ।(স্পেশালাইজড রাইবোসোম হাইপোথিসিস)। [৬৯][৭০] তবে এই অনুকল্প নিয়ে যথেষ্ট বিতর্ক রয়েছে এবং এ বিষয়টি নিয়ে গবেষণা চলছে।[৭১][৭২] রাইবোসোমের গঠনের ভিন্নতা প্রোটিন সংশ্লেষণ নিয়ন্ত্রণে জড়িত থাকে এমনটি প্রথম প্রস্তাব করেন বিজ্ঞানী ভিন্স মাউরো এবং জেরাল্ড এডেলম্যান।[৭৩] তারা রাইবোসোমের নিয়ন্ত্রণ কার্যাবলী ব্যাখ্যা করার জন্য রাইবোসোম ফিল্টার হাইপোথিসিস প্রস্তাব করেছিলেন। প্রমাণসমূহ দেখাচ্ছে বিভিন্ন কোষ জনসংখ্যার জন্য নির্দিষ্ট বিশেষ রাইবোসোম জিনের ট্রান্সলেশনে কিভাবে প্রভাব রাখে ।[৭০] কিছু রাইবোসোমাল প্রোটিন সাইটোসোলিক কপির[৭৪] সম্মিলিত কমপ্লেক্স থেকে বিনিময় করে যা নির্দেশ করে ইন ভিভো রাইবোসোমের গঠন সম্পূর্ণ নতুন রাইবোসোম সংশ্লেষ না করেই পরিবর্তন করা যেতে পারে।কিছু রাইবোসোমাল প্রোটিন সেলুলার জীবনের জন্য অত্যাবশ্যকীয় , কিছু ততটা গুরুত্বপূর্ণ নয়। [৭৫] উদীয়মান খামিরে, ১৪/৭৮ রাইবোসোমাল প্রোটিনগুলি বৃদ্ধির জন্য অপ্রয়োজনীয়, যেখানে মানুষের মধ্যে এটি কোষের উপর নির্ভর করে। ভিন্নধর্মীতার অন্যান্য রূপের মধ্যে রয়েছে রাইবোসোমাল প্রোটিনের সংশ্লেষণ-পরবর্তী পরিবর্তন যেমন অ্যাসিটাইলেশন, মিথিলেশন এবং ফসফোরাইলেশন।[৭৬] অ্যারাবিডোপসিস[৭৭][৭৮][৭৯][৮০] ভাইরাসের অভ্যন্তরীণ রাইবোসোম এন্ট্রি সাইট (IRESs) গঠনগতভাবে স্বতন্ত্র রাইবোসোম দ্বারা সংশ্লেষণের মধ্যস্থতা করতে পারে। উদাহরণস্বরূপ, খামির এবং স্তন্যপায়ীদের কোষে eS২৫ ছাড়া ৪০S রাইবোসোমাল ইউনিট CrPV IGR IRES গ্রহণ করতে অক্ষম।[৮১]
রাইবোসোমাল আরএনএ এর বিভিন্নতা কাঠামোগত ভারসাম্যে এবং কার্যাবলিতে গুরুত্বপূর্ণ ভূমিকা পালন করে এবং বেশিরভাগ mRNA এর পরিবর্তনগুলি অত্যন্ত সংরক্ষিত অঞ্চলে পাওয়া যায়।[৮২][৮৩] সবচেয়ে সাধারণ আর আরএনএর পরিবর্তনগুলি হল সিউডোরিডিলেশন এবং রাইবোজের ২’-O মিথিলেশন।[৮৪]
লুই পাস্তুর (ফরাসি: Louis Pasteur লুই পাস্ত্যোর্) (ডিসেম্বর ২৭, ১৮২২ – সেপ্টেম্বর ২৮, ১৮৯৫) একজন ফরাসিঅণুজীববিজ্ঞানী ও রসায়নবিদ।[২] তিনিই প্রথম আবিষ্কার করেন যে অণুজীব অ্যালকোহলজাতীয় পানীয়ের পচনের জন্য দায়ী। জীবাণুতত্ত্ব ও বিভিন্ন রোগ নির্মূলে বিভিন্ন ধরনের প্রতিষেধক আবিষ্কার করে স্মরণীয় হয়ে রয়েছেন।
প্রারম্ভিক জীবন
লুই পাস্তুর ১৮২২ সালের ২৭ ডিসেম্বর ফ্রান্সের জুরা প্রদেশের দোল শহরে জন্মগ্রহণ করেন ও আরবোয়া শহরে বেড়ে উঠেন।[৩] দরিদ্র পিতা সেখানকার একটি ট্যানারিতে চাকুরি করতেন। ১৮৪৭ সালে পাস্তুর ফ্রান্সের একোল থেকে পদার্থবিদ্যা ও রসায়নে ডক্টরেট ডিগ্রি অর্জন করেন। সেখানে তিনি জৈব যৌগেরআলোক সমাণুতা নিয়ে গবেষণা করেন। তিনি দেখান যে, আলো যখন জৈব যৌগের দ্রবনের ভেতর দিয়ে যায় তখন এর দিক পরিবর্তন ঘটে। তিনি প্রস্তাব করেন যে, একই জৈব যৌগ যাদের গঠন এক, তারা সমাণু হতে পারে যদি তারা একে-অপরের আলো প্রতিবিম্ব হয়।
১৮৪৮ সালে দিজোঁ লিসিতে সংক্ষিপ্ত সময়ের জন্য রসায়নের অধ্যাপক হিসেবে চাকরি করেন। সেখানেই বিশ্ববিদ্যালয়ের রেক্টরের কন্যা মারি লরেন্তের সাথে প্রণয়ে আবদ্ধ হন। ২৯ মে, ১৮৪৯ সালে তারা বিবাহ-বন্ধনে আবদ্ধ হন। পাঁচ সন্তানের তিনটিই প্রাপ্তবয়স্ক হবার পূর্বেই টাইফয়েড রোগে মারা যায়। ব্যক্তিগতভাবে নির্মম এ ঘটনায় তিনি মুষড়ে না পড়ে এর প্রতিকারে মনোনিবেশ ঘটিয়েছিলেন।
কর্মজীবন
তিনি গবেষণা কর্ম চালিয়ে যান এবং সেই সাথে দিজোঁ ও স্ত্রাসবুর্গ বিশ্ববিদ্যালয়ে শিক্ষকতা শুরু করেন। ১৮৫৪ সালে পাস্তুর স্থানীয় এক বিশ্ববিদ্যালয়ে বিজ্ঞান বিভাগের ডীন হিসেবে নিয়োগ লাভ করেন। তখন থেকেই তিনি স্থানীয় মদের কলগুলোতে গাঁজন প্রক্রিয়া নিয়ে গবেষণা শুরু করেন। তিনি দেখান অ্যালকোহল উৎপাদন ইস্টের পরিমানের উপর নির্ভর করে। তিনি আরও প্রমাণ করেন মদের অম্লতা তাতে ব্যাক্টেরিয়ার ক্রিয়ার জন্য ঘটে।
গবেষণা
মদ শিল্প
মদেরঅম্লতা ফ্রান্সের মদ ব্যবসাতে এক বিশাল সমস্যা ছিল। এর ফলে প্রতিবছর অনেক অর্থ গচ্চা যেত। পাস্তুর মদের স্বাদ ঠিক রেখে ব্যাক্টেরিয়া মুক্ত করার জন্য গবেষণা শুরু করেন। তিনি দেখেন মদকে গরম করলে ব্যাক্টেরিয়া মরে যায় এবং মদের কোন পরিবর্তন হয় না। পাস্তুর একই পদ্ধতি দুধের ক্ষেত্রেও প্রয়োগ করেন এবং ভাল ফল পান। পাস্তুরের এই পদ্ধতি বিশ্বব্যাপী এখন ব্যবহৃত হচ্ছে। তার নামানুসারে এই পদ্ধতিকে পাস্তুরায়ন নামে নামাঙ্কিত করা হয়।
জীবনের উৎপত্তি
পাস্তুর এখন মদে ব্যক্টেরিয়ার উৎস নিয়ে গবেষণা শুরু করেন। তৎকালীন সময়ে অনেকে ধারণা করতেন ব্যাক্টেরিয়া নির্জীব বস্তু থেকে আপনা আপনি সৃষ্টি হয়। এর বিপক্ষেও অনেকে বিজ্ঞানী ছিলেন। গ্রিক দার্শনিকএরিস্টটলের সময় থেকেই এই বিতর্ক ছিল, কিন্তু কোন বিজ্ঞানসম্মত উত্তর ছিল না। পাস্তুর পরীক্ষার মাধ্যমে দেখান, নির্জীব বস্তু থেকে ব্যাক্টেরিয়া বা কোন রকম জীবনের সূত্রপাত হতে পারে না। তিনি প্রমাণ করেন, মদে বাতাস ও অন্যান্য মাধ্যম থেকে ব্যাক্টেরিয়া আসে। পাস্তুর তার বিখ্যাত পরীক্ষার সাহায্যে যেটা দেখিয়েছিলেন তা হল, জীবাণুমুক্ত নিয়ন্ত্রিত (পাস্তুর প্রদত্ত) পরিবেশে প্রাণ আপনা আপনি জন্ম নেয় না; কিন্তু অন্য পরিবেশে অন্য ভাবে যে কখনই জন্ম নিতে পারবে না – এই কথা কিন্তু পাস্তুরের ফলাফল হলফ করে বলেনি
পাস্তুরের পরীক্ষা স্বতঃজননবাদকে (Theory of spontaneous generation) ভুল প্রমাণ করেছে। স্বতঃজননতত্ত্বের দাবীদারদের সবাই বিশ্বাস করতেন জটিল জীব তার পূর্ণ অবয়বে নিজে নিজেই ‘সৃষ্টি’ হয়। যেমন, এরিস্টটল বিশ্বাস করতেন কিছু মাছ এবং পতংগের মত ছোট প্রাণী স্বতঃস্ফুর্তভাবে উদ্ভূত হয় । ব্রিটিশ গবেষক আলেকজান্দার নীডহ্যাম (১১৫৭-১২১৭) বিশ্বাস করতেন ফার গাছ সমুদ্রের লবণাক্ত পানিতে ফেলে রাখলে তা থেকে রাজহাঁস জন্ম নেয়। জ্যান ব্যাপটিস্ট হেলমন্ট (১৫৮০-১৬৪৪) ভাবতেন ঘর্মাক্ত নোংরা অন্তর্বাস ঘরের কোনায় ফেলে রাখলে তা থেকে ইঁদুর আপনা আপনিই জন্ম নেয়। বিজ্ঞানী পুশে (১৮০০-১৮৭২) বিশ্বাস করতেন খড়ের নির্যাস থেকে ব্যাকটেরিয়া বা অণুজীব স্বতঃস্ফুর্তভাবেই জন্ম নেয়[2]। পাস্তুরের গবেষণা মূলতঃ এই ধরনের ‘সৃষ্টিবাদী’ ধারণাকেই বাতিল করে দেয়। কিন্তু পাস্তুরের পরীক্ষা কিংবা জৈবজনির কোন সূত্রই বলে না যে, প্রাথমিক জীবন জড় পদার্থ থেকে তৈরি হতে পারবে না
রেশম শিল্প
১৮৬৫ সালে ফ্রান্স সরকার পাস্তুরকে ফ্রান্স রেশম শিল্পের সমস্যা সমাধানে আহ্বান জানায়। এক মহামারীতেরেশম পোকার উৎপাদন হ্রাস পেয়েছিল। পাস্তুর দেখেন রেশম পোকার এই সমস্যা বংশগত এবং মায়ের থেকে পরবর্তী প্রজন্মে সংক্রামিত হতে পারে। তিনি প্রস্তাব করেন কেবলমাত্র রোগ মুক্ত গুটি বাছাই করার মাধ্যমেই রেশম শিল্পকে বাঁচানো সম্ভব।
জীবাণু তত্ত্ব
পাস্তুর দেখান কিছু রোগ অণুজীব দ্বারা সংঘটিত হতে পারে, যারা জল ও বাতাসের মাধ্যমে ছড়ায়। তিনি তার জীবাণু তত্ত্বে দেখান যে অণুজীব বৃহদাকার জীবকে আক্রমণ করে রোগ সংঘটিত করতে পারে।
টিকা আবিষ্কার
অ্যান্থ্রাক্স
পাস্তুর প্রথম অ্যানথ্রাক্সেরটিকা আবিষ্কার করেন। তিনি গবেষণার মাধ্যমে বুঝতে পারেন গৃহপালিত পশুতে অ্যান্থ্রাক্স ব্যাসিলি (Bacillus anthrasis)-এর আক্রমণেই অ্যান্থ্রাক্স হয়। তিনি রোগ সৃষ্টিতে অক্ষম অ্যান্থ্রাক্স ব্যাসিলি ভেড়ায়ইনজেকসনের মাধ্যমে প্রবেশ করান এবং দেখেন পরবর্তীকালে এগুলো আর রোগ সৃষ্টিতে সক্ষম ব্যাসিলি দিয়ে আক্রান্ত হয় না।
জলাতঙ্ক
অ্যান্থ্রাক্স প্রতিরোধক আবিস্কারের পর পাস্তুর অন্যান্য রোগের প্রতিরোধের জন্য গবেষণা শুরু করেন। তিনি জলাতঙ্ক নিয়ে কাজ করে দেখেন এটি নার্ভাস সিস্টেমের একটি রোগ এবং আক্রান্ত পশুর স্পাইনাল কর্ডের নির্যাস দ্বারা অন্য প্রাণিকে জলাতঙ্কে আক্রান্ত করা যায়। এই পদ্ধতিতে তিনি রোগ প্রতিরোধে অক্ষম জলাতঙ্ক ভাইরাস উৎপাদন করেন, যা জলাতঙ্কের টিকা হিসেবে ব্যবহার করা সম্ভব। ১৮৮৫ সালে পাস্তুর প্রথম এক শিশু বালকের উপর এই টিকা প্রয়োগ করেন। ছেলেটিকে জলাতঙ্ক আক্রান্ত কুকুর কামড়িয়েছিল, তারপর ছেলেটির মা তাকে পাস্তুরের গবেষণাগারে নিয়ে আসেন। পাস্তুর ছেলেটিকে টিকা প্রদান করেন এবং ছেলেটি ভাল হয়ে উঠে।
মৃত্যু
জলাতঙ্কের টিকা আবিস্কারের পরে ফ্রান্স সরকার পাস্তুর ইনস্টিটিউট স্থাপন করেন। এই ইনস্টিটিউটের পরিচালক থাকাকালীন ১৮৯৫ সালে লুই পাস্তুর মৃত্যুবরণ করেন।[৪]
ত্বক হচ্ছে মেরুদন্ডী প্রাণীর বহিরাঙ্গিক একটি অংশ যা প্রকৃতপক্ষে একটি নরম আবরণ এবং দেহকে আবৃত করে রাখে। এটি প্রাণিদের ভিতরের অংশগুলোকে রক্ষা করে। এটি মানবদেহের সবচেয়ে বড় অঙ্গ। স্তন্যপায়ীদের ত্বকে লোম বা ছোট চুল থাকে। ত্বক পারিপার্শ্বিক পরিবেশের সাথে সংযোগ স্থাপন করে এবং বাহ্যিক প্রভাবের বিরুদ্ধে এটি দেহের প্রাথমিক রক্ষক। রোগ সৃষ্টিকারী জীবাণুর বিরুদ্ধে ও শরীর থেকে অতিরিক্ত পানি অপসারণ প্রতিরোধে ত্বক গুরুত্বপূর্ণ ভূমিকা রাখে। এছাড়া অন্তরক, তাপমাত্রা নিয়ন্ত্রক, ইন্দ্রিয় ও ভিটামিন ডি উৎপাদক হিসেবে ত্বক কাজ করে।
ত্বকের পুরুত্ব প্রাণীভেদে এবং একই প্রাণীতে অবস্থানভেদে পরিবর্তিত হয়। মানুষের চোখের পাতার ত্বক সবচেয়ে পাতলা আর হাত-পায়ের তালুর ত্বক সবচেয়ে পুরু।
স্তরঃ
ত্বকের স্তর প্রধানত দুটি। এপিডার্মিস (বাইরে) ও ডার্মিস (ভিতরে)। এপিডার্মিস পাতলা আর ডার্মিস পুরু।
এপিডার্মিস ৫টি স্তর নিয়ে গঠিত। ১ কঠিন স্তর (stratum corneum)
এবং বৃদ্ধির প্রয়োজনে জীব (উদ্ভিদ এবং প্রাণী) খাদ্য গ্রহণ করে । খাদ্যদ্রব্যগুলোর পরিপাক এবং বিপাক এর কালে কিছু অপ্রয়োজনীয় পদার্থ সৃষ্টি করে । অপ্রয়োজনীয় এবং ক্ষতিকর বস্তুগুলো একটি বিশেষ প্রক্রিয়ায় দেহ থেকে নিষ্কাষিত হয় । প্রক্রিয়াটির নাম রেচন।
নিঃসরণ বা বহিঃষ্করণ এবং রেচন এক নয় । নিঃসরণে বস্তুসমূহ কোষ নির্গত হওয়ার পর নির্দিষ্ট কাজ সম্পন্ন করে । রেচন প্রক্রিয়ায় নিষ্কাষিত বস্তুগুলোকে বলা হয় রেচন পদার্থ । রেচন পদার্থগুলো সাধারণত ক্ষারীয় হয় । রেচন প্রক্রিয়ায় অংশগ্রহণকারী অঙ্গগুলোকে বলা হয় রেচন অঙ্গ ।
এককোষী জীবসমূহে রেচন পদার্থসমূহ সরাসরি কোষের বাইরে নির্গত হয় ।
জলজ প্রাণী সমূহ বর্জ্য পদার্থ হিসেবে সাধারনত অ্যামোনিয়া ত্যাগ করে । অ্যামোনিয়া উচ্চমাত্রায় দ্রবণীয় বলে পানিতে এটা দ্রবীভূত থাকে ।
পাখিরা নাইট্রোজেনজাত বর্জ্য পদার্থ ইউরিক অ্যাসিড ত্যাগ করে ।
ইনসেকটা পর্বের প্রাণীদের ম্যালপিজিয়ান নালিকা নামক রেচন অঙ্গ থাকে । ম্যালপিজিয়ান নালিকা রেচন পদার্থ সমূহ দেহের বাইরে ত্যাগ করে । কিছু রেচন পদার্থ মালপিজিয়ান নালিকায় আজীবনের জন্য সঞ্চিত থাকে ।
উদ্ভিদের রেচনঃ
প্রাণীদের তুলনায় উদ্ভিদের বিপাক ক্রিয়া ধীরগতিতে সম্পন্ন হয় এবং উদ্ভিদের নির্দিষ্ট রেচন অঙ্গ না থাকায় বর্জ্য পদার্থের নিষ্কাশন ও একটি ধীর প্রক্রিয়া ।
সবুজ উদ্ভিদ এবং ক্লোরোফিলবিহীন উদ্ভিদ অন্ধকারে শ্বসন এর সময় বর্জ্য পদার্থ হিসেবে কার্বন ডাই অক্সাইড এবং জল উৎপন্ন করে । কার্বন ডাই অক্সাইড দিনের বেলা সালোকসংশ্লেষণে ব্যবহৃত হয়ে যায়। সালোকসংশ্লেষণে উৎপন্ন অক্সিজেনস্টোমাটা, আন্তঃকোষীয় ছিদ্র ইত্যাদির মাধ্যমে বের করে দেওয়া হয় । উদ্ভিদের অতিরিক্ত পানি প্রস্বেদন এর মাধ্যমে বের করে দেওয়া হয় । গাছের ঝরে যাওয়া পাতাতেও কিছু রেচন পদার্থ সঞ্চিত থাকতে দেখা যায়।
একটি লিম্ফোব্লাস্ট হলো পরিবর্তিত সেল মরফোলজি সহ একটি পরিবর্তিত নিষ্পাপ লিম্ফোসাইট y। এটি ঘটে যখন লিম্ফোসাইট একটি অ্যান্টিজেন দ্বারা সক্রিয় করা হয় (অ্যান্টিজেন-উপস্থাপক কোষ থেকে) এবং নিউক্লিয়াস এবং সাইটোপ্লাজম বৃদ্ধি দ্বারা নতুন এমআরএনএ এবং প্রোটিন সংশ্লেষণ দ্বারা পরিমাণে বৃদ্ধি পায়। তারপরে লিম্ফোব্লাস্ট ৩-৫ দিনের জন্য প্রতি ২৪ ঘণ্টা থেকে ৪ বার বিভাজন শুরু করে, একটি একক লিম্ফোব্লাস্টের সাথে প্রায় ১ হাজার ক্লোন তৈরি করে তার মূল মত লিম্ফোসাইট, প্রতিটি ভাগের সাথে অনন্য অ্যান্টিজেনের বৈশিষ্ট্য ভাগ করে নেওয়া হয়। অবশেষে বিভাজনকারী কোষগুলি ইফেক্টর কোষগুলিতে পৃথক হয়, যাকে প্লাজমা সেল (বি কোষের জন্য), সাইটোঅক্সিক টি কোষ এবং হেল্পার টি কোষ হিসাবে পরিচিত। [১]
লিম্ফোব্লাস্টগুলি অপরিপক্ব কোষগুলিকেও উল্লেখ করতে পারে যা সাধারণত পরিপক্ব লিম্ফোসাইটগুলি গঠনে পৃথক করে। [২] সাধারণত অস্থি মজ্জার মধ্যে লিম্ফোব্লাস্টগুলি পাওয়া যায় তবে তীব্র লিম্ফোব্লাস্টিক লিউকেমিয়া (ALL) এ লিম্ফোব্লাস্টগুলি অনিয়ন্ত্রিতভাবে প্রসারিত হয় এবং পেরিফেরিয়াল রক্তে প্রচুর সংখ্যায় পাওয়া যায়।
যদিও লিম্ফোব্লাস্ট সাধারণত লিউকোসাইটগুলির পরিপক্বতায় পূর্ববর্তী কোষকে বোঝায়, এই শব্দটির ব্যবহার কখনও কখনও অসঙ্গত হয়। ক্রনিক লিম্ফোসাইটিক লিউকেমিয়া রিসার্চ কনসোর্টিয়াম একটি লিম্ফোব্লাস্টকে সংজ্ঞায়িত করে “একটি লিম্ফোসাইট যা অ্যান্টিজেন দ্বারা উদ্দীপিত হওয়ার পরে আরও বড় হয়ে উঠেছে। লিম্ফোব্লাস্টগুলি অপরিপক্ব লিম্ফোসাইটগুলির মতো দেখায় এবং একসময় পূর্ববর্তী কোষ বলে মনে করা হত। । [৪] সাধারণত, লিউকেমিয়া সম্পর্কে কথা বলার সময়, “ব্লাস্ট” লিম্ফোব্লাস্টগুলির সংক্ষেপণ হিসাবে ব্যবহৃত হয়।
লিম্ফোব্লাস্টগুলি মাইলোব্লাস্ট থেকে মাইক্রোস্কোপিকভাবে আলাদা আলাদা নিউকোলিওলি, আরও ঘনীভূত ক্রোমাটিন এবং সাইটোপ্লাজমিক গ্রানুলের
অনুপুস্থিতিতেদ্বারা চিহ্নিত করা যায়। তবে এই মরফোলজিক পার্থক্যগুলি নিখুঁত নয় এবং একটি নির্দিষ্ট রোগ নির্ণয় পৃথকীকরণের রিসেপ্টরগুলির অনন্য ক্লাস্টারের উপস্থিতির জন্য অ্যান্টিবডি ইমিউনস্টেইনিংয়ের উপর নির্ভর করে। [৫]
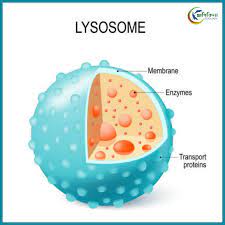
লাইসোসোম বা লাইসোজোম (ইংরেজি:Lysosome) এক ধরনের কোষীয় অঙ্গাণু যা সাধারণত প্রাণী কোষে পাওয়া যায়।কোষের সাইটোপ্লাজমে দ্বি-স্তর বিশিষ্ট লিপো-প্রোটিন সম্বনয়ে গঠিত মেমব্রন বা ঝিল্লি দ্বারা আবৃত যে অঙ্গাণুটি নানাবিধ হাইড্রোলাইটিক এনজাইমের ধারক বা বাহক হিসেবে কাজ করে তাই লাইসোজোম বলে।এতে বিদ্যমান ভেসিকলগুলো হাইড্রোলাইটিক এনজাইম এর আধার হিসেবে কাজ করে। একে “আত্মঘাতী থলিকা” বা “আত্মঘাতী স্কোয়াড” বলা হয়। লাইসোসোম শব্দটি গ্রিক থেকে আগত। গ্রিক “লাইসো” শব্দের অর্থ হজমকারী এবং “সোমা” শব্দের অর্থ বস্তু।
বেলজিয়ামের ক্যাথলিক ইউনিভার্সিটি অফ লুভ এর অধ্যাপক ক্রিশ্চিয়ান দে দুভ এবং তার দল ১৯৪৯ সালে যকৃতেইনসুলিনের ক্রিয়া-কৌশল নিয়ে গবেষণা চালাচ্ছিলেন। এসময় তারা শর্করা বিপাকের (গ্লাইকোলাইসিস) ক্ষেত্রে একটি গুরুত্বপূর্ণ এনজাইম এবং ইনসুলিনের লক্ষ্যবস্তু গ্লুকোজ ৬-ফসফাটেজ (G6P) এনজাইমের ওপর লক্ষ্য রেখেছিলেন। তারা দেখতে পেলেন যে, রক্তে শর্করার মাত্রা নিয়ন্ত্রণের প্রধান এনজাইম হল G6P।[১][২] কিন্তু বারবার পরীক্ষার পরও তারা কোষীয় নির্যাস থেকে এনজাইমটি আলাদা করতে পারেন নি। তাই এনজাইমটি আলাদা করতে তারা আরও cell fractionation পদ্ধতি অবলম্বন করেন।[৩]
এনজাইমটি সণাক্ত করতে তারা অ্যাসিড ফসফাটেজ ব্যবহার করে একটি পদ্ধতি অবলম্বন করেন। কিন্তু এর ফলাফল ছিল অপ্রত্যাশিতভাবে খুবই কম; প্রত্যাশিত মানের ১০% এর কাছাকাছি। এরপর একদিন তারা ৫ দিন ধরে সংরক্ষিত কিছু বিশুদ্ধ কোষ ভগ্নাংশের ক্রিয়াকলাপ পরিমাপ করেন। তারা লক্ষ্য করেন, এনজাইমের ক্রিয়া বৃদ্ধি পেয়ে আবার নতুন নমুনাসমূহের মাত্রায় উন্নীত হয়েছে। পরীক্ষাটি বারবার করেও একই রকম ফলাফল পাওয়া যায়। এ কারণে প্রস্তাব করা হয় যে, কোন একটি প্রতিবন্ধকতার জন্য ভিত্তিস্তরে এনজাইমের দ্রুত প্রবেশ সীমাবদ্ধ থাকে। ফলে একটি নির্দিষ্ট সময় পার হবার পরই কেবল এনজাইমগুলোর ব্যাপন ঘটে। তারা এই প্রতিবন্ধকতাকে “ঝিল্লি দ্বারা পরিবেষ্টিত ও অ্যাসিড ফসফাটেজ ধারণকারী একটি থলের ন্যায় কাঠামো ”- বলে আখ্যা দেন।[৪][৫]
ঝিল্লিযুক্ত ভগ্নাংশ, যেগুলো কোষীয় অঙ্গাণু বলে পরিচিত, সেখান থেকে একটি সম্পর্কহীন এনজাইম cell fractionation প্রক্রিয়ার সাথে পাওয়া যায়। এদের পরিপাকীয় বৈশিষ্ট্যের সাথে মিল রেখে, ১৯৫৫ সালে দে দুভ এদের নাম দেন “লাইসোসোম”।[৬]
ভারমন্ট বিশ্ববিদ্যালয় এর অ্যালেক্স বি. নভিকফ সেই বছরই দে দুভ এর গবেষণাগার পরিদর্শনে যান এবং ইলেকট্রন মাইক্রোস্কোপ ব্যবহার করে সফলভাবে লাইসোসোম অঙ্গাণুটির প্রথম দৃশ্যমান প্রমাণ সংগ্রহ করেন। এছাড়া অ্যাসিড ফসফাটেজের জন্য একটি রঞ্জক পদ্ধতি ব্যবহার করে, দে দুভ ও নভিকফ লাইসোসোমের পানিগ্রাহী এনজাইমের অবস্থান নিশ্চিত করেন।[৭]
অ্যালেক্স বি. নভিকফ ১৯৬০ সালে এদের আণুবীক্ষণিক গঠন বর্ণনা করেন। ১৯৬০ সালে Matile এটি নিউরোস্পোরা ছত্রাকে পর্যবেক্ষণ করেন।
লাইসোসোম দ্বি-স্তর বিশিষ্ট লিপোপ্রোটিন আবরণী দ্বারা আবদ্ধ থাকে। ভ্যাকুওল ঘন তরলপূর্ণ ও এসিডিক হয়। লাইসোসোম সাধারণত বৃত্তকার। এর ব্যাস সাধারণত ০.২ থেকে ০.৮ মাইক্রোমিলি এর মধ্যে হয়।[৯] তবে এদের আকার ছোট-বড় হতে পারে। যেমন: বৃক্ক কোষের লাইসোসোম অপেক্ষাকৃত বড় হয়। এমন কি বড় লাইসোসোম এর আকার ছোটগুলোর চেয়ে ১০ গুণেরও বেশি বড় হতে পারে।
লাইসোসোমে ৬০ টিরও বেশি এনজাইম এবং ঝিল্লিতে ৫০টিরও বেশি ধরনের মেমব্রেন প্রোটিন পাওয়া গেছে।[১১][১২] এ এনজাইমগুলো অমসৃণ এন্ডোপ্লাজমিক রেটিকুলাম এ সংশ্লেষিত হয়। প্রধান এনজাইমগুলো হল:
১.প্রাথমিক লাইসোসোম: প্রাথমিক সদ্য গঠিত এনজাইম পূর্ণ ক্ষুদ্র থলি বিশেষ।
২.অটোফ্যাগোসোম বা সাইটোলাইসোসোম: কোষের অপ্রয়োজনীয় অবাঞ্ছিত বা ক্ষতিগ্রস্ত অংশ মসৃণ পর্দা দ্বারা আবৃত হয়ে পৃথক অংশ গঠন করে। এর সাথে এক বা একাধিক প্রাথমিক লাইসোসোম মিলিত হয়ে অটোফ্যাগোসোম গঠন করে। এতে মাইটোকন্ড্রিয়া, রাইবোসোম, পারঅক্সিসোম, গ্লাইকোজেন দানা ইত্যাদির স্বপাচন ঘটে।
৩.হেটারোফ্যাগোসোম বা গৌণ লাইসোসোম:ফ্যাগোসাইটোসিস বা পিনোসাইটোসিসের ফলে উৎপন্ন ফ্যাগোসোমের সাথে একাধিক প্রাথমিক লাইসোসোম মিলিত হয়ে হেটারোফ্যাগোসোম গঠন করে।
৪.রেসিডুয়াল বডি বা টেলো লাইসোসোম: অপাচিত বস্তু যুক্ত গৌণ লাইসোসোমকে টেলো লাইসোসোম বলে।
৫, উদ্ভিদ কোষে সম্প্রতি একস্তর বিশিষ্ট Oleosome নামে লাইসোসোম পাওয়া গেছে ।
কাজ
ফ্যাগোসাইটোসিস ও পিনোসাইটোসিস প্রক্রিয়ায় জীবাণু ধ্বংস করে।
প্রোটিনকে ডাইপেপটাইড ও কার্বোহাইড্রেটকে মনোস্যাকারাইডে পরিণত করে।
কোষে খাদ্যাভাব দেখা দিলে লাইসোসোমের এনজাইমগুলো কোষের অঙ্গাণুগুলো ধ্বংস করে দেয়। একে স্বগ্রাস বা অটোফ্যাগি বলে। এভাবে সমগ্র কোষ পরিপাক হয়ে গেলে তাকে অটোলাইসিস বলে।
লাইসোসোমের এনজাইমগুলো কোষের মধ্যে গৃহীত খাদ্যবস্তু, ব্যাকটেরিয়া ইত্যাদি জারিত করলে তাকে হেটারোফ্যাগি বলে।
কোষ বিভাজনকালে এরা কোষীয় ও নিউক্লিও আবরণী ভাঙতে সহায়তা করে। এরা কোষে কেরাটিন প্রস্তুত করে।
লাইসোসোম কোষের ক্ষতি করে না কেন?
লাইসোসোমের ঝিল্লি কোষের বাকি অংশকে ক্ষতিগ্রস্ত হওয়া থেকে রক্ষা করে।
লাইসোসোমের অভ্যন্তর এসিডিক। এর পি.এইচ প্রায় ৪.৫ থেকে ৫ এর মধ্যে [১৪] এনজাইমগুলো পি.এইচ সেনসেটিভ এবং শুধু এসিডিয় পরিবেশে কাজ করে। সাইটোসল (সাইটোপ্লাজম এর ম্যাট্রিক্স) এর পি.এইচ (৭.২) প্রশম বলে কোষের অ্যালকালাইন পরিবেশে এরা কাজ করে না।
রোগ
লাইসোসোমে এনজাইমের ঘাটতির ফলে কিছু পদার্থ (যেমন: গ্লাইকোজেন, গ্লাইকোলিপিড) লাইসোসোমে সঞ্চিত যা ২০ প্রকারের কনিজেনিট্যাল রোগ সৃষ্টি করে। এদের লাইসোসোমাল স্টোরেজ ডিজিস (Lysosomal storage disease) বলে। এর সরাসরি কোনো চিকিৎসা নেই।[১৫]
Lysomal Storage Diseases ব্লাড ক্যান্সার এর ক্ষেত্রে নিউক্লিয়েজ এনজাইম নিউক্লিয়াসে জিনগত পরিবর্তন ঘটায়
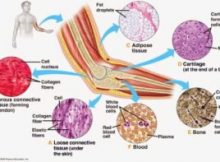
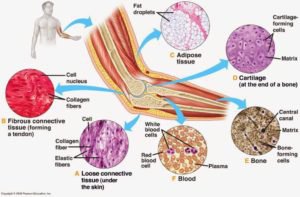
যোজক কলা এক ধরনের আঁশবহুল কলা, যেটি মানবদেহের অন্যান্য অঙ্গ অথবা কলাকে সমর্থন করে, যুক্ত করে অথবা বিচ্ছিন্ন করে। এটি চার ধরনের কোষকলার একটি[১] (অপর তিনটি হচ্ছে আবরণী, পেশি ও স্নায়ুকলা)।
যোজক কলা ফাইব্রোব্লাস্ট, এডিপোসাইট, ম্যাক্রোফেজ, মাস্ট কোষ এবং লিউকোসাইড নিয়ে গঠিত। ১৮৩০ সালে জোসেফ পিটার মুলার “যোজক কলা” শব্দটি (জার্মান ভাষায়, বিন্দেগিউবে) প্রস্তাব করেন। ১৮ শতকেই যোজক কলা স্বতন্ত্র্য শ্রেণি হিসেবে বিবেচিত হত। প্রাণীদের যোজক কলার মূল উপাদান কোলাজেন ও স্তন্যপায়ী প্রাণীদের মধ্যে এটির পরিমাণ সবচেয়ে বেশি, যা প্রায় ২৫%।
যোজক কলার সাধারণ বৈশিষ্ট্যঃ ১.কোষগুলো সাধারণত নির্দিষ্ট স্তরে বিন্যস্ত থাকে না; ২.কোষ গুলোর মাঝে ম্যাট্রিক্স থাকায় ঘন সন্নিবেশিত হয় না; ৩.মেসোডার্ম নামক ভ্রুন থেকে এ কলার উৎপত্তি; ৪.রক্তবাহ উপস্থি।
সাধারণ কার্যাবলীঃ
১.দেহের বিভিন্ন অঙ্গ বা কলার মধ্যে যোগসূত্র সৃষ্টি করে; ২.কঙ্কাল গঠন করে দেহের বিভিন্ন কাঠামো প্রদান এবং বিভিন্ন অঙ্গের সংরক্ষণে চলাচলে সাহায্য করে; ৩.বিভিন্ন পদার্থের পরিবহনে অংশগ্রহণ করে; ৪.দেহের উপবিষ্ট ক্ষতিকারক পদার্থ ও জীবণু থেকে রক্ষা করে; ৫.ক্ষত নিরাময়ের অংশগ্রহণ করে; ৬.দেহের তাপ নিয়ন্ত্রণ করে। [২][৩]
তরল যোজক কলা
যে কলা ধাত্র তরল ও যার কোষগুলো ধাত্রে ভাসমান অবস্থায় থাকে, তাকে তরল যোজক কলা বলে। এটি শোষিত খাদ্যসহ বিভিন্ন দ্রবীভূত পদার্থ দেহের বিভিন্ন অংশে পরিবহন করে এবং রেচন পদার্থ ধৌত করে বলে একে পরিবহন বা সংবহন কলাও বলা হয়। তরল যোজক কলা প্রধানত দুই প্রকার। যথা:
যৌন দ্বিরূপতা (ইংরেজি: Sexual dimorphism) বলতে কোন নির্দিষ্ট প্রজাতির জীবের স্ত্রী ও পুরুষ সদস্যের মধ্যে বাহ্যিক শারীরিক বা চারিত্রিক বৈশিষ্ট্যের এমন তারতম্য দেখা যায় যাতে করে স্ত্রী-পুরুষে খুব সহজে পার্থক্য করা যায়। সাধারণত প্রধান পার্থক্যটি হল যৌনাঙ্গের বিভিন্নতা। এছাড়া বর্ণ, আকার-আকৃতি, গঠন অথবা কোন বিশেষ জিনগত বৈশিষ্ট্যের মাধ্যমে এদের আলাদা করা যায়। প্রধানত দুটো কারণে যৌন দ্বিরূপতা দেখা যায়। প্রথমত, বিপরীত লিঙ্গের জীবকে আকৃষ্ট করার লক্ষে যৌন বিবর্তনের মাধ্যমে দ্বিরূপতা সৃষ্টি (যেমন পুরুষ ময়ূরের ঝলমলে পালক) এবং দ্বিতীয়ত, প্রতিরক্ষার উদ্দেশ্যে দ্বিরূপতা সৃষ্টি (যেমন পুরুষ বেবুনের বড় দেহ ও শ্বদন্ত)। প্রধানত পাখিদের মধ্যে যৌন দ্বিরূপতা বেশি পরিলক্ষিত হয়। পুরুষ পাখিদের ঝলমলে ও উজ্জ্বল পালক থাকে, এতে প্রজনন ও সীমানা বজায় রাখতে সুবিধা হয়। স্ত্রী পাখিদের পালক সাধারণত খুব সাদামাটা হয়, যাতে প্রাকৃতিক পরিবেশের সাথে তা একদম মিশে যায়। ফলে বাসায় বসে থাকা স্ত্রী পাখিরা শত্রুর হাত থেকে বেঁচে যায়। একই কারণে স্তন্যপায়ী ও অমেরুদণ্ডী প্রাণীদের মধ্যে যৌন দ্বিরূপতা দেখা যায়। এছাড়া আচরণের দিক থেকেও বিভিন্নতা দেখা যায়। যেমন: কাঁটাওয়ালা তক্ষকের পুরুষ সদস্যদের খাদ্যাভ্যাস স্ত্রী সদস্যদের তুলনায় ভিন্ন।[১]